| Data | Observed | Predicted | (RE) | Unit | Description | Reference |
|---|
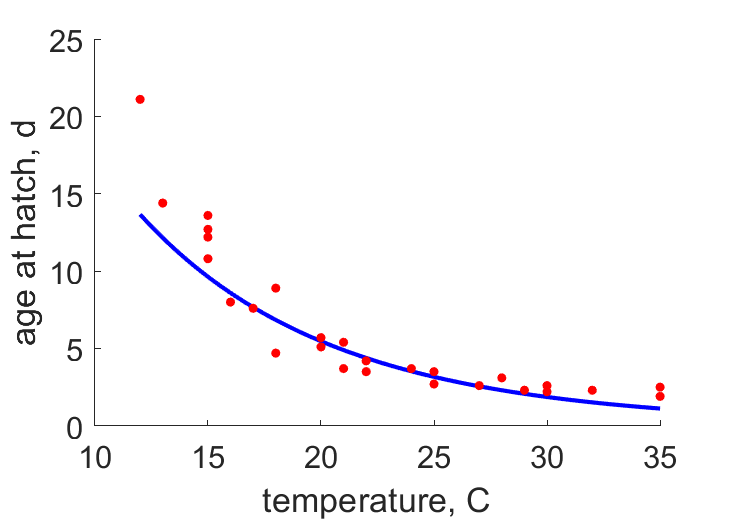
| ahT21 | 5 | 4.896 | (0.0208) | d | age at hatch | Sund1966 |
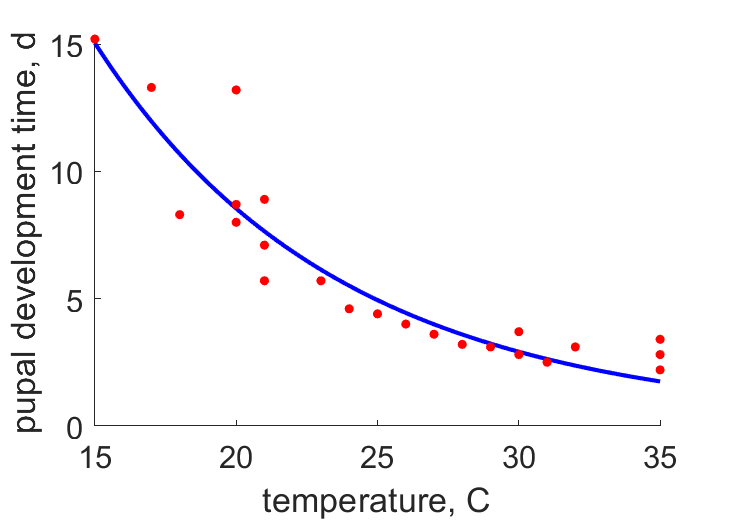
| teT21 | 8.5 | 7.644 | (0.1007) | d | time since pupation at emergence | Sund1966 |
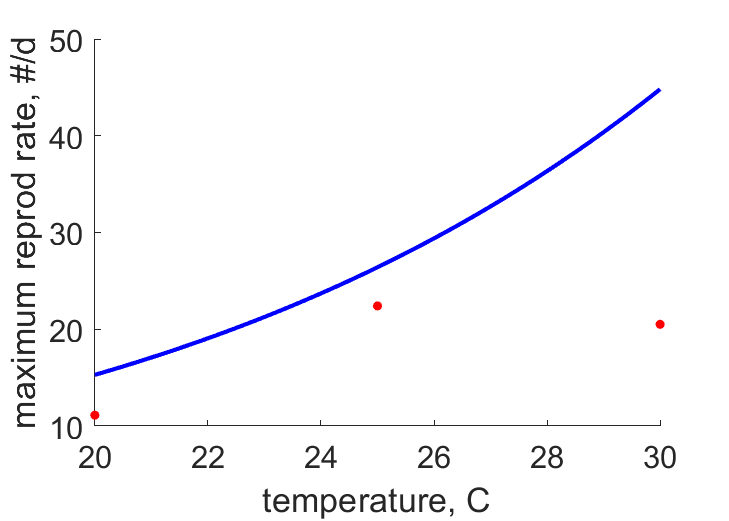
| RiT21 | 12.52 | 20.04 | (0.6005) | #/d | maximum reprod rate | Sund1966 |
| ahT25 | 2.42 | 3.165 | (0.3078) | d | age at hatch | OmkaSriv2003 |
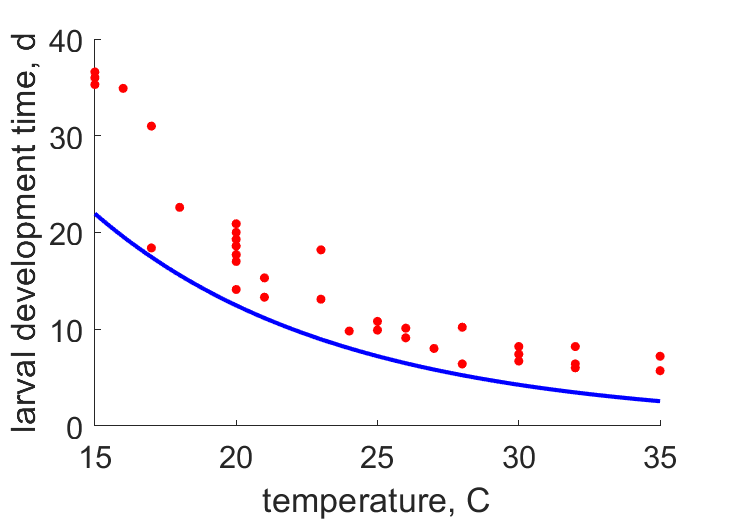
| t1T25 | 1.72 | 1.753 | (0.01938) | d | duration of instar 1 | OmkaSriv2003 |
| t2T25 | 1.64 | 1.545 | (0.05789) | d | duration of instar 2 | OmkaSriv2003 |
| t3T25 | 1.84 | 1.697 | (0.07761) | d | duration of instar 3 | OmkaSriv2003 |
| t4T25 | 2.75 | 2.211 | (0.1959) | d | duration of instar 4 | OmkaSriv2003 |
| teT25 | 3.55 | 4.941 | (0.3918) | d | time since pupation at emergence | OmkaSriv2003 |
| tR125 | 8.2 | 6.625 | (0.1921) | d | pre-oviposition period | OmkaSriv2003 |
| RiT25 | 25.27 | 31.01 | (0.2268) | #/d | maximum reprod rate | OmkaSriv2003 |
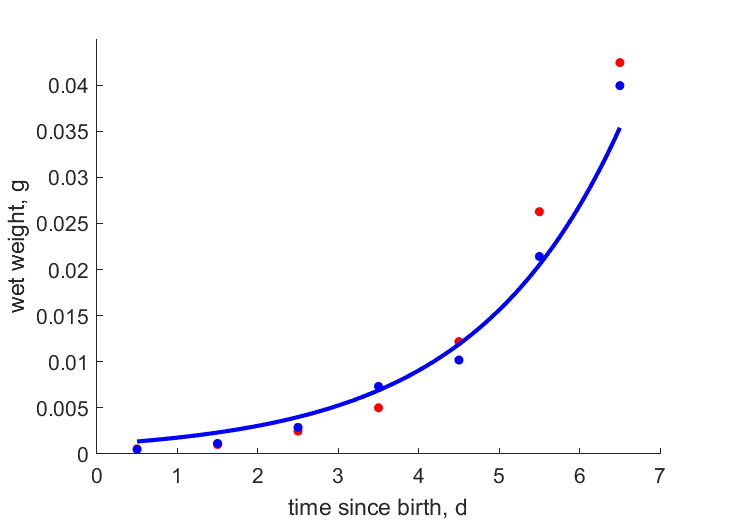
| Ww1T25 | 0.00226 | 0.001841 | (0.1855) | g | wet weight of instar 1 | OmkaSriv2003 |
| Ww2T25 | 0.00555 | 0.004417 | (0.2041) | g | wet weight of instar 2 | OmkaSriv2003 |
| Ww3T25 | 0.01063 | 0.01088 | (0.02321) | g | wet weight of instar 3 | OmkaSriv2003 |
| Ww4T25 | 0.01979 | 0.0338 | (0.7078) | g | wet weight of instar 4 | OmkaSriv2003 |
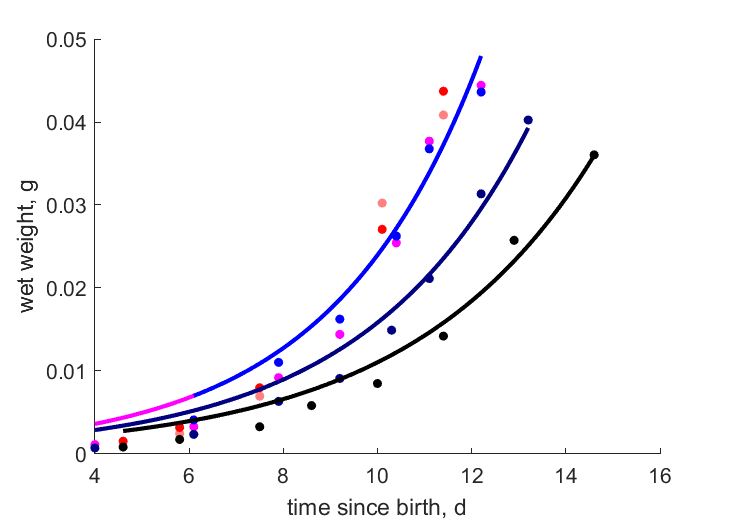
| WwjT25 | 0.01505 | 0.05202 | (2.456) | g | wet weight of pupa | OmkaSriv2003 |
| WweT25 | 0.04063 | 0.03344 | (0.177) | g | wet weight of female imago after emergence | OmkaSriv2003 |
| ahT23 | 3.8 | 3.931 | (0.03435) | d | age at hatch | ObryOrr1990 |
| t1T23 | 3.1 | 2.178 | (0.2976) | d | duration of instar 1 | ObryOrr1990 |
| t2T23 | 2.2 | 1.919 | (0.1278) | d | duration of instar 2 | ObryOrr1990 |
| t3T23 | 2.4 | 2.108 | (0.1217) | d | duration of instar 3 | ObryOrr1990 |
| t4T23 | 5.4 | 2.746 | (0.4914) | d | duration of instar 4 | ObryOrr1990 |
| teT23 | 6.3 | 6.136 | (0.02596) | d | time since pupation at emergence | ObryOrr1990 |
| WweT23 | 0.0377 | 0.03344 | (0.1131) | g | wet weight of female imago after emergence | ObryOrr1990 |
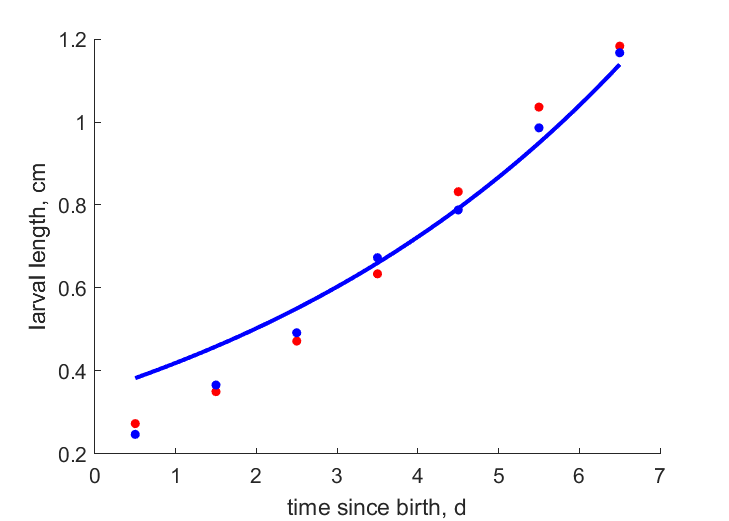
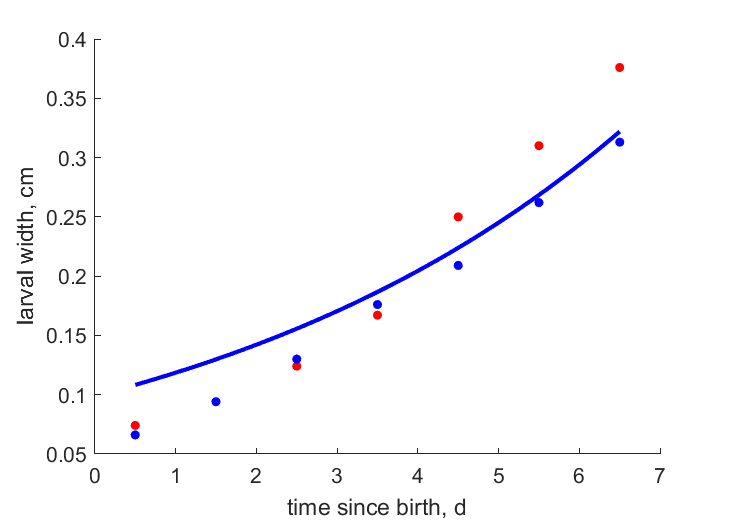
| L1eT23 | 0.765 | 0.7655 | (0.0007008) | cm | body length of female imago after emergence | ObryOrr1990 |
| L2eT23 | 0.591 | 0.5711 | (0.03364) | cm | body width female imago after emergence | ObryOrr1990 |
| ahT28 | 3 | 2.299 | (0.2337) | d | age at hatch | SingSing1993 |
| t1T28 | 1.76 | 1.274 | (0.2763) | d | duration of instar 1 | SingSing1993 |
| t2T28 | 1.8 | 1.122 | (0.3765) | d | duration of instar 2 | SingSing1993 |
| t3T28 | 2.4 | 1.233 | (0.4863) | d | duration of instar 3 | SingSing1993 |
| t4T28 | 3.15 | 1.606 | (0.4901) | d | duration of instar 4 | SingSing1993 |
| teT28 | 6.68 | 3.589 | (0.4627) | d | time since pupation at emergence | SingSing1993 |
| tR128 | 6.66 | 4.812 | (0.2774) | d | pre-oviposition period | SingSing1993 |
| RiT28 | 47.62 | 42.69 | (0.1036) | #/d | maximum reprod rate | SingSing1993 |
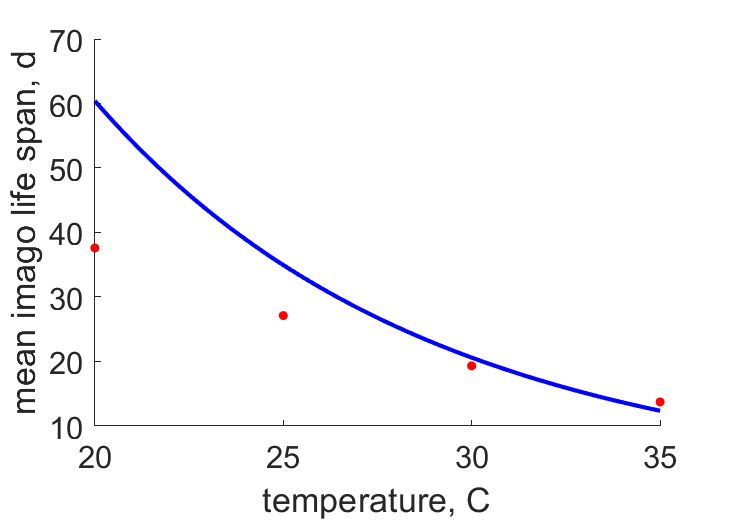
| aemT28 | 26.74 | 24.5 | (0.08395) | d | life span of female imago | SingSing1993 |
| ahT24 | 3.56 | 3.526 | (0.009648) | d | age at hatch | FaroZhu2020 |
| t1T24 | 2 | 1.953 | (0.02337) | d | duration of instar 1 | FaroZhu2020 |
| t2T24 | 2 | 1.721 | (0.1394) | d | duration of instar 2 | FaroZhu2020 |
| t3T24 | 2.47 | 1.891 | (0.2345) | d | duration of instar 3 | FaroZhu2020 |
| t4T24 | 2.29 | 2.463 | (0.07575) | d | duration of instar 4 | FaroZhu2020 |
| teT24 | 4.17 | 5.504 | (0.32) | d | time since pupation at emergence | FaroZhu2020 |
| tR124 | 3.26 | 7.38 | (1.264) | d | pre-oviposition period | FaroZhu2020 |
| RiT24 | 30.21 | 27.83 | (0.07864) | #/d | maximum reprod rate | FaroZhu2020 |
| aemT24 | 48.16 | 37.57 | (0.22) | d | life span of female imago | FaroZhu2020 |
| pA1T27 | 2.502e+04 | 1.858e+04 | (0.2574) | J/g.d | dry-weight-specific assimilation rate at instar 1 | Sing1976 |
| pA2T27 | 2.983e+04 | 1.858e+04 | (0.3772) | J/g.d | dry-weight-specific assimilation rate at instar 2 | Sing1976 |
| pA3T27 | 3.72e+04 | 1.858e+04 | (0.5005) | J/g.d | dry-weight-specific assimilation rate at instar 3 | Sing1976 |
| pA4T27 | 3.732e+04 | 1.858e+04 | (0.5021) | J/g.d | dry-weight-specific assimilation rate at instar 4 | Sing1976 |
| pAeT27 | 4.916e+04 | 3.496e+04 | (0.2888) | J/g.d | dry-weight-specific assimilation rate of imago | Sing1976 |
| tjT25 | 8.9 | 7.207 | (0.1902) | d | time since hatch at pupation | KaluHode2004 |
| WweT252 | 0.0388 | 0.03344 | (0.1382) | g | wet weight of female imago after emergence | KaluHode2004 |
| Ww0 | 0.000187 | 0.001213 | (5.485) | g | initial wet weight of an egg | ChatNaz2020 |
| Wwj | 0.0424 | 0.05202 | (0.2268) | g | wet weight of fresh pupa | Baye2024 |
| Ww14d | 0.039 | 0.03344 | (0.1426) | g | wet weight of female imago after emergence (14 d old) | Baye2024 |
| Ww21d | 0.0659 | 0.0692 | (0.05006) | g | wet weight of older female imago (21 d old) | Baye2024 |
| L114d | 0.769 | 0.7655 | (0.004504) | cm | body length of female imago after emergence (14 d old) | Baye2024 |
| L214d | 0.588 | 0.5711 | (0.02871) | cm | body width of female imago after emergence (14 d old) | Baye2024 |
| Wwj2 | 0.0415 | 0.05202 | (0.2534) | g | wet weight of fresh pupa | Baye2024 |
| Ww14d2 | 0.0359 | 0.03344 | (0.06861) | g | wet weight of female imago after emergence (14 d old) | Baye2024 |
| Ww24d | 0.0658 | 0.0692 | (0.05166) | g | wet weight of older female imago (24 d old) | Baye2024 |
| L114d2 | 0.72 | 0.7655 | (0.06324) | cm | body length of female imago after emergence (14 d old) | Baye2024 |
| L214d2 | 0.552 | 0.5711 | (0.03463) | cm | body width of female imago after emergence (14 d old) | Baye2024 |
| L124d | 0.756 | 0.7655 | (0.01261) | cm | body length of female imago after emergence (24 d old) | Baye2024 |
| L224d | 0.555 | 0.5711 | (0.02904) | cm | body width of female imago after emergence (24 d old) | Baye2024 |