Predictions & Data for this entry
| Model: asj | climate: MC | migrate: | phylum: |
| COMPLETE = 3.0 | ecozone: MA | food: biPz | class: |
| MRE = 0.213 | habitat: 0iMp | gender: Hh | order: |
| SMSE = 0.134 | embryo: Mp | reprod: O | family: |
Zero-variate data
| Data | Observed | Predicted | (RE) | Unit | Description | Reference |
|---|---|---|---|---|---|---|
| ab | 1 | 1.174 | (0.1736) | d | age at birth | JaspHara2013 |
| tp | 13 | 16.48 | (0.2675) | d | time since birth at puberty | BakeReev1974 |
| am | 244 | 243.8 | (0.0008614) | d | life span | issg |
| Lb | 0.04 | 0.06133 | (0.5333) | cm | oral-aboral length at birth | Mart1987 |
| Lj | 1 | 1.132 | (0.1317) | cm | oral-aboral length at metam | RapoNova2005 |
| Lp | 3 | 4.253 | (0.4178) | cm | total length at puberty | ReevSyms1989 |
| Li | 6 | 5.531 | (0.07815) | cm | ultimate oral-aboral length | WalrLang2013 |
| Wc0 | 0.22 | 0.1115 | (0.4933) | mug | initial carbon weight of egg | JaspCost2014 |
| Wwi | 50 | 52.09 | (0.04176) | g | ultimate wet weight | Jasp2012, WalrLang2013 |
| R78 | 1.123e+04 | 4469 | (0.6021) | #/d | reprod rate at 78 mm total length | JaspCost2014 |
Uni- and bivariate data
| Data | Figure | Independent variable | Dependent variable | (RE) | Reference |
|---|---|---|---|---|---|
| LR | 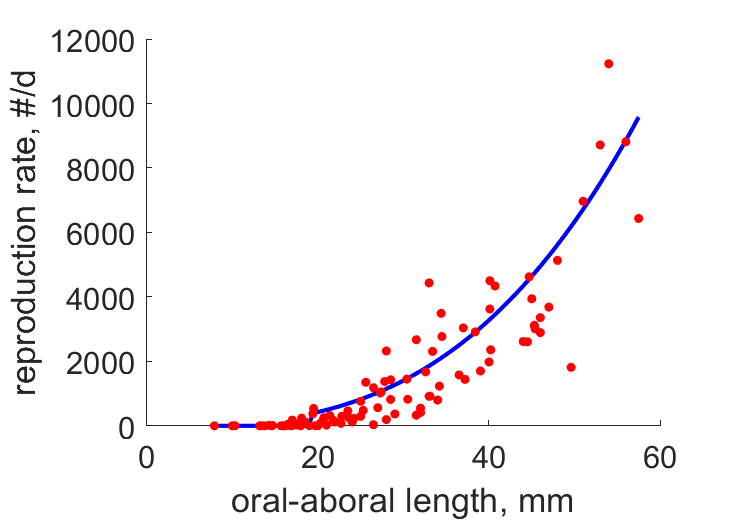 | oral-aboral length | reproduction rate | (0.4518) | Jasp2012 |
| LR2 | 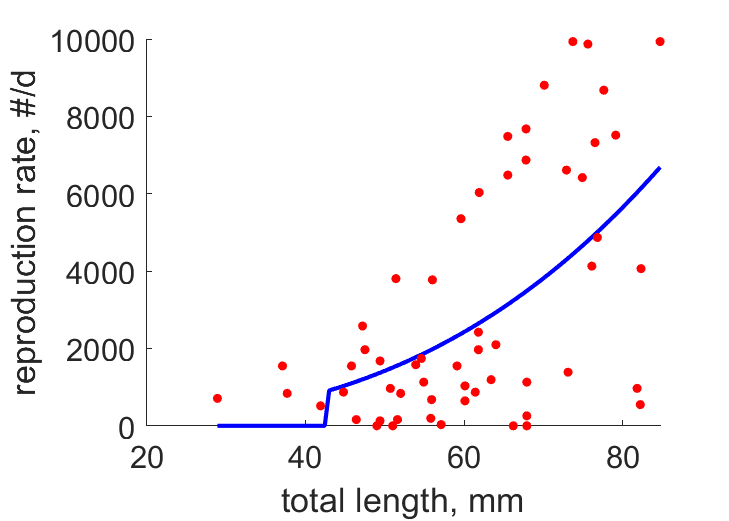 | total length | reproduction rate | (0.6704) | BakeReev1974 |
| LV | 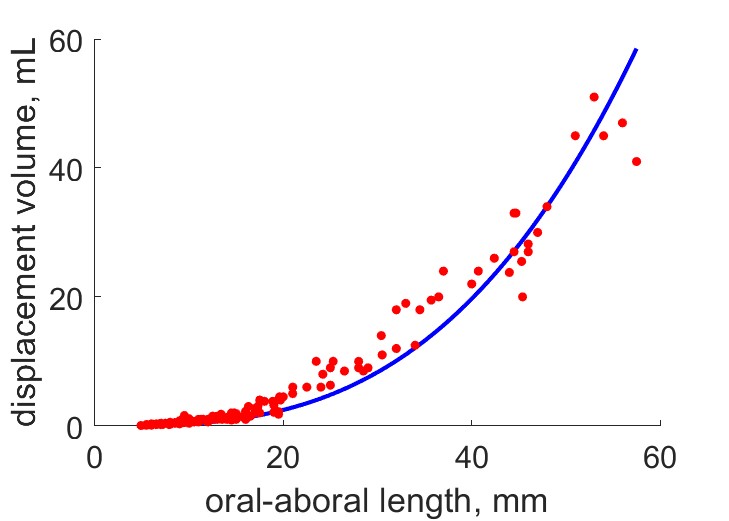 | oral-aboral length | displacement volume | (0.2289) | Jasp2012 |
| LWC | 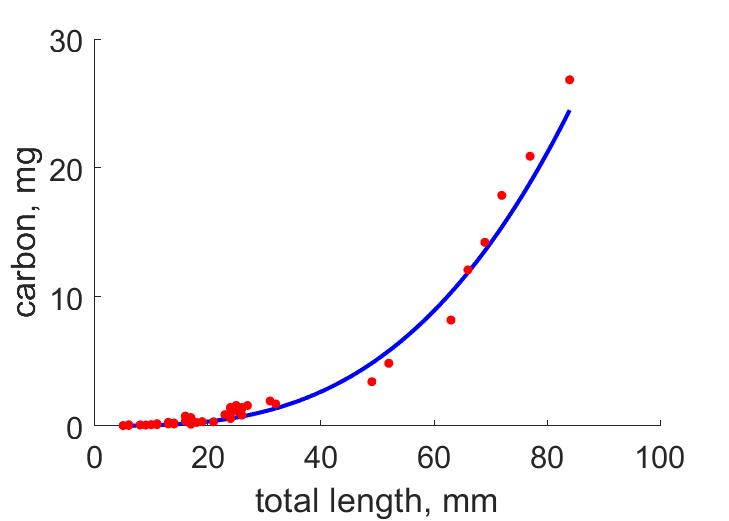 | total length | carbon | (0.1746) | ReevSyms1989 |
| LWw | 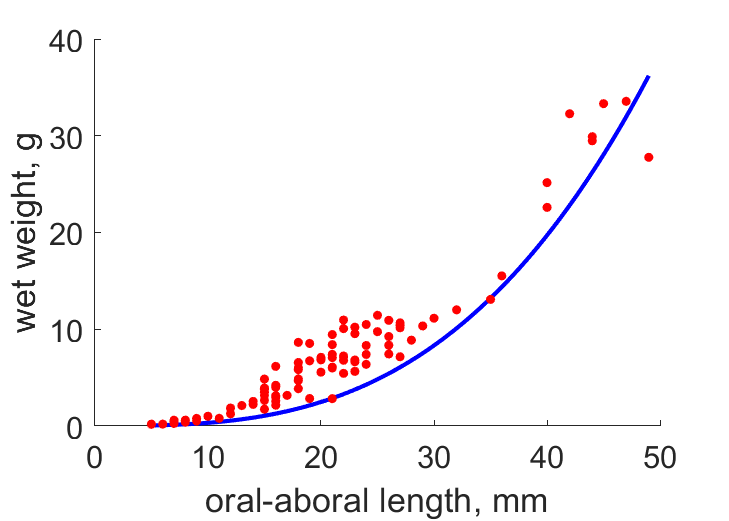 | oral-aboral length | wet weight | (0.398) | WalrLang2013 |
| WwR | 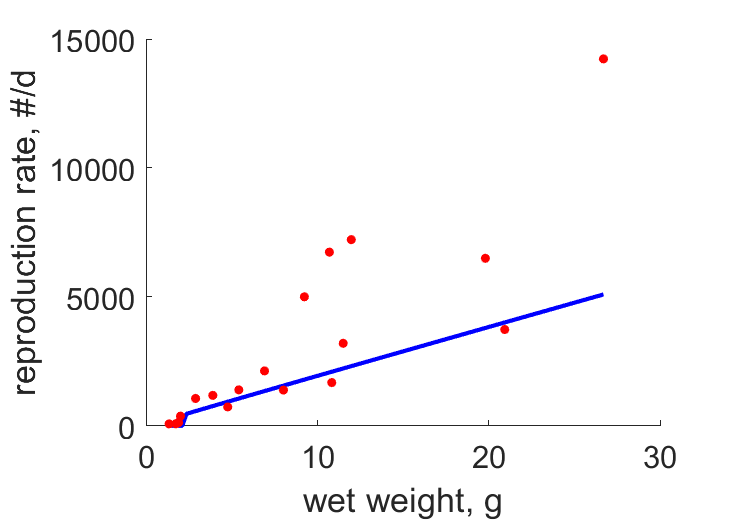 | wet weight | reproduction rate | (0.5163) | Krem1976 |
| tL |  | time since birth | total length | (0.1278) | BakeReev1974 |
| tN | 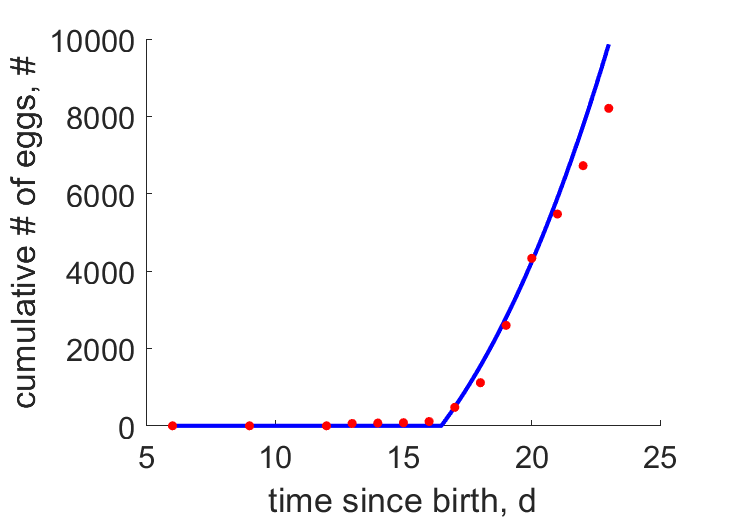 | time since birth | cumulative # of eggs | (0.1425) | BakeReev1974 |
Pseudo-data at Tref = 20°C
| Data | Generalised animal | Mnemiopsis leidyi | Unit | Description |
|---|---|---|---|---|
| v | 0.02 | 0.03153 | cm/d | energy conductance |
| p_M | 18 | 109.2 | J/d.cm^3 | vol-spec som maint |
| k_J | 0.002 | 0.002 | 1/d | maturity maint rate coefficient |
| k | 0.3 | 0.001439 | - | maintenance ratio |
| kap | 0.8 | 0.3911 | - | allocation fraction to soma |
| kap_G | 0.8 | 0.7989 | - | growth efficiency |
| kap_R | 0.95 | 0.475 | - | reproduction efficiency |
Discussion
- Length measurement are either: [1] total length= including lobes (TL), [2] oral-aboral length, i.e. aboral pole to mouth (OA) or [3] oral-statocyst length, i.e. statocyst to mouth opening (OS). But there are more types of length measurements, see Mutl1999
- The type of length measurement is not specified in BakeReev1974 (data from Fig.1, Tab2) nor in ReevSyms1989. Pers. comm. with Particia Kremer tells that it is total length (including the lobes). The length measurement is also really sensitive to whether or not the animal was immerged in water (Pers. Comm. P. Kremer)
- There might be the possibility for larvae to reproduce sexually while they are still larvae, a condition known as dissogeny but which was only observed in LAB (and in M. mccrady by Mart1987)
- metamorphosis (as defined by switching from V1 morphic growth to isomorphic growth) is assumed to occur at end of cydippid larval stage
- LR-data were given weight zero because zero-reprod data distort estimation
Facts
- This entry is discussed in AuguJasp2014 (Ref: AuguJasp2014)
- Data from all Mnemiopsis leidyi and mccradyi species can be combined as the genus is considered monospecific until further research is done (Ref: Bayh2005)
- There are no benthic resting stages. Mnemiopsis sp. has 3 larval stages: tentaculate (from hatching to ca. 4 mm), transition (ca. 5 - 10 mm) and lobate (from about 6 - 18 mm) distinctly different morphologies, which can influence diet composition, feeding rates and digestion. (Ref: RapoNova2005)
- After the cydippid larvae stage (= tentaculate) the tentacles disappear and the lobes are formed. Diet switches from microzooplankton to mesozooplankton (0.2-20 mm) (Ref: RapoNova2005)
- The species is a simultaneous hermaphrodite able to perform self-fertilisation (Ref: nobanis)
Bibliography