Concentration-response versus biology-based methods
The present EU-project Nomiracle succeeds a previous EU-project
Mixtox, which resulted a.o. in an Excel spreadsheet for the analysis
of toxicity data for mixtures of compounds. We naturally applied this
spreadsheet to our data on the effects of
mixtures of metals on the survival of Folsomia candida. Jan
Baas found that the outcome of the spreadsheet is sensitive to the
type of baseline model that is selected (concentration addition
versus independent action), and for each of the two possible
choices the type of interaction changed several times during the 21 d
exposure. With our biology-based methods we
found no interaction between the metals, which strongly suggests that
these inconsistent outcomes are just artifacts.
On invitation by Hans Løkke and David Spurgeon, I discussed in the
Nomiracle meeting at Antwerp why the spreadsheet produces problematic
results. This is because it is based on concentration-response curves
of the log-logistic family (including e.g. log-probit, gamma and
Weibull models), and this whole family is problematic for modelling
effects on survival. The reasons are in the assumptions behind these
models:
- individuals have identical toxico-kinetics
- they die for sure if the internal concentration exceeds some
threshold
- threshold values vary among individuals and follow some
frequency distribution
Three compelling reasons why these assumptions cannot be true are
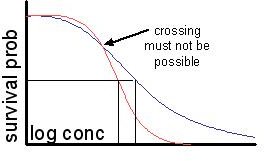 the slope of the
concentration response curve typically becomes steeper during
exposure. If the assumptions were true, it should remain
constant. Even for descriptive purposes the curves are problematic,
because as soon as the
slopes are allowed to vary in time, curves for different exposure
times will intersect, with the implication that the survival
probability increases in time for some concentrations, which is not
possible. The change in slope during exposure is well-captured
by the biology-based method, which links hazard rates linearly to
internal concentrations; it is caused by toxico-kinetics.
the slope of the
concentration response curve typically becomes steeper during
exposure. If the assumptions were true, it should remain
constant. Even for descriptive purposes the curves are problematic,
because as soon as the
slopes are allowed to vary in time, curves for different exposure
times will intersect, with the implication that the survival
probability increases in time for some concentrations, which is not
possible. The change in slope during exposure is well-captured
by the biology-based method, which links hazard rates linearly to
internal concentrations; it is caused by toxico-kinetics.- experiments indicate that re-exposed cohorts have the same LC50
value as the first-exposed ones (Newman & Closkey, 2000}, while we
would expect that it is higher if the assumptions would be true,
because sensitive individuals are removed by the first
exposure.
- sublethal effects don't support large
physiological differences among individuals. If reproduction is
affected, it does so in all individuals. The assumptions imply that
the reason why not all individuals die at the same moment is because
of these differences.
Notice that goodness-of-fit arguments are not among these
reasons! But just selecting another descriptive curve, goodness-of-fit
could easily be improved, but the above-mentioned problems will remain.
Concentration-response curves for lethal effects are, therefore, not
realistic. They are also problematic in eco-toxicity applications. This
is because
- the incorporation of exposure time is problematic for the reasons
that are already discussed; LC50's and small-effect concentrations
depend on exposure time.
- small-effect concentrations are difficult to determine and
model-sensitive because of the practical difficulty to find enough
concentrations that cause partial effects. The selection of the
model is rather arbitrary.
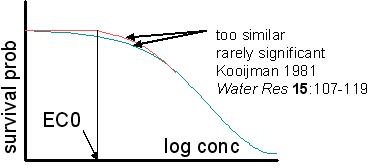 the curves are incompatible with the NOEC. Attempts to solve
this problem by the
incorporation of a NEC in these models turned out to be less
practical, because the resulting NEC value rarely differs from zero
significantly. The biology-based models don't suffer from this
problem, because they analyse all exposure times simultaneously and
are typically found to be significantly different from zero.
the curves are incompatible with the NOEC. Attempts to solve
this problem by the
incorporation of a NEC in these models turned out to be less
practical, because the resulting NEC value rarely differs from zero
significantly. The biology-based models don't suffer from this
problem, because they analyse all exposure times simultaneously and
are typically found to be significantly different from zero.- the results are difficult to extrapolate from individuals to
populations, from one chemical or species of organism to another.
- the method is not applicable to time-varying concentrations
- the method gives misleading results for mixtures
The arguments for sublethal effects are only slightly different. These
are all strong reasons
to cease the application of the concentration-response methods,
especially because the much more powerful biology-based methods are
available.
Contrary to biology-based methods, concentration-addition and
independent-action are different concepts for concentration-response
methods for mixtures of compounds. Since the slopes of any pair of
compounds are typically different (and change in time), concentration
addition is really weird for this method, and leads to complex
implicit survival probabilities as functions of the (external)
concentration of the compounds. No justification is known. For
biology-based methods, however, concentration-addition comes naturally
as a consequence of independent action. If changes in the internal
concentration can be neglected, the effect on the survival probability
is via the product of concentration and exposure time; this is well
known in pharmacology, and in some practical application of toxicants
(for instance in disinfection of buildings). This further underpins
the need for the retirement of concentration-response methods.
In Antwerp, I briefly discussed the alternatives that the
biology-based methods offer and how they can be used for extrapolation
pruposes. JRC-ECB will organise a workshop in 2007 on biology-based
methods for environmental risk assessment, for which Tjalling Jager
and I will provide the content. We will organise a DEB tele-course in
2007. The free registration for this course is already open.
Look here for
futher illustrations
Go to projects' entry