Drs. D. (Daniel) Bontje

Predicting Unexpected Effects of Chemicals on Ecosystems
Introduction
Standard bioassays on the potential of chemicals for biological
effects focus on acute or chronic survival, growth and reproduction of
single species under constant optimal physiological laboratory
conditions. The effects of a chemical depend on its concentration, its
mode of action, the choice of test species and the exposure time. Due
to absence of interactions among organisms (food competition,
predation, crowding) and (nutrient) recycling, it is difficult to
translate these single-species effects to a community. The variability
of field conditions, which affect a.o. the bio-aviability, further
contribute to the problem of translating effects from single-species
bioassays to effects on communities in the field. Interactions among
biota and feedback loops via nutrient availability make that it might
be difficult even to recognize a deviating behaviour of a community as
an effect of a toxic substance.
Aim
The first aim of this project is to understand the effect on a
community, given observed effects on the species that occur in this
community for low concentrations of a number of key chemicals
(i.e. industrial chemicals that are frequently released in the
environment, and have different modes of action). To this end we use a
simple microcosm consisting of a decomposer, a producer, a consumer
and a predator (which eats both producer and consumer). The role of
the producers is to fix energy (light) and inorganic carbon for
metabolic work, of the decomposers to unlock nutrients that are locked
in organic compounds and of the consumers and the predators to speed
up the recycling of nutrients.
Canonical community
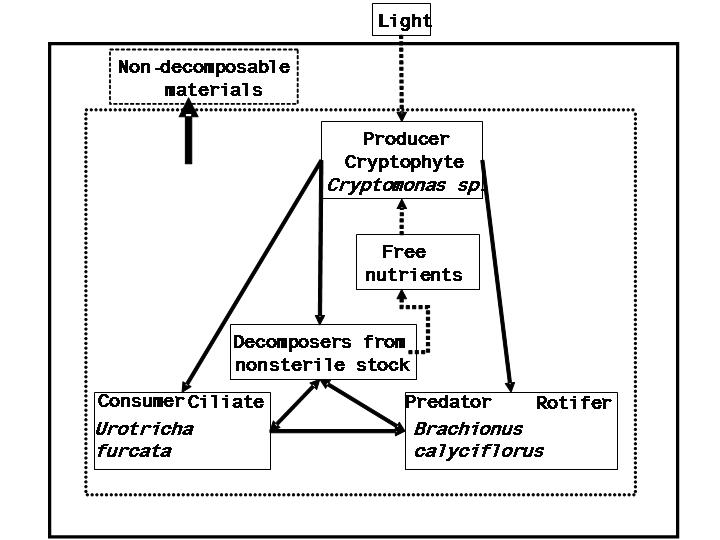
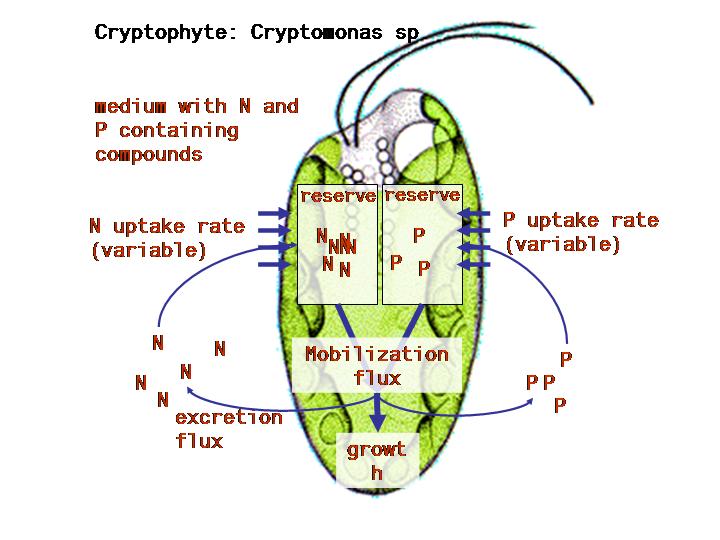 We choose the single species for each category of organism and call
this simple system a canonical community as within this community all
basic activities are found that are present in a larger ecosystem. For
the role of producer we choose the mixotrophic cryptophyte
Cryptomonas spec.; it has proven to be an excellent food source
for several species of ciliates and can easily remain in suspension
with help of its flagella. The consumer will be the planktonic ciliate
Urotricha furcata, which is abundant in many stratifying
temperate lakes. It feeds on both bacteria and cryptophytes, with a
preference for the latter. The predator will be the rotifer
Brachionus calyciflorus, which is also unbundant in many
freshwater systems. It has a relatively short generation cycle and was
used in many studies on feeding behavior and predator/prey
relationships. It is able to consume cryptophytes, ciliates and even
bacteria. The decomposers will be bacteria that originate from
non-axenic ciliate stocks cultures; their species composition will not
be determined.
We choose the single species for each category of organism and call
this simple system a canonical community as within this community all
basic activities are found that are present in a larger ecosystem. For
the role of producer we choose the mixotrophic cryptophyte
Cryptomonas spec.; it has proven to be an excellent food source
for several species of ciliates and can easily remain in suspension
with help of its flagella. The consumer will be the planktonic ciliate
Urotricha furcata, which is abundant in many stratifying
temperate lakes. It feeds on both bacteria and cryptophytes, with a
preference for the latter. The predator will be the rotifer
Brachionus calyciflorus, which is also unbundant in many
freshwater systems. It has a relatively short generation cycle and was
used in many studies on feeding behavior and predator/prey
relationships. It is able to consume cryptophytes, ciliates and even
bacteria. The decomposers will be bacteria that originate from
non-axenic ciliate stocks cultures; their species composition will not
be determined.
We try to simplify further and select nutrient regimes where only
nitrogen is limiting. Thus we give our test-organisms in the microcosm
a constant amount of light and temperature and a surplus of all other
resources.
Biofilms
As a second choice for a simplified community, we will study biofilms
in this project, in close collaboration with ms. dr. Mechthild
Schmitt-Jansen (UFZ) and ms. Christina Klünder (UFZ) on Modelkey SITE
3. The system has many aspects in common with the canonical community
in terms of the biota and their interactions, but differs in having
more species, a spatial structure and an open (nutrient) exchange with
their surroundings. Moreover it has the possibility to expose biofilms
under field conditions.
Determination of the normal behavior
All organisms need at least carbon, oxygen, nitrogen and phosphorous
molecules and energy to build new biomass and to maintain and repair
themselves. Homeostasis (i.e. the ability to have a relatively
constant body composition) comes with stoichiometric constraints on
community dynamics. Chemicals can influence the uptake rates, and all
other metabolic rates and efficiencies. To be able to determine the
effects of the chemicals on the canonical community we first need to
know the normal behavior of this community, and understand this
behaviour in terms of eco-physiological properties of species. E.g.:
what is the growth rate of Cryptomonas given a certain nutrient
level. How many Cryptomonas can maximal be eaten per Urotricha per
day? How many Cryptomonas are there needed to make one new Urotricha?
Given an initial nutrient level how much of each organism will there
be after a few weeks? Does cyclic behavior occur? (Models frequently
predict cyclic behaviour at high nutrient levels, a phenomenon known
as the paradox of enrichment.)
Modeling approach
We use the Dynamic Energy Budget
theory to quantify the eco-physiological behaviour of the species
and to predict the behaviour of the canonical community, given
parameter estimates that are derived from single-species experiments.
The toxicity module of this
theory captures effects of chemical compounds as changes of parameter
values as functions of the internal concentration of compounds. The
modes of action of the compound determines which parameters are
effected; this can be species-specific and can depend on the
concentration. The compound can e.g. increase the maintenance
requirement or the costs of growth, or decrease the (maximum)
nutrient/food uptake rate.
By comparing the deviating behavior with the expected normal behavior
we will be able to say which variables (amounts of nutrients and of
the various biota, turnover rates) have changed in the exposed
canonical community or biofilm. To this end, the parameter values
needed to feed the model will be estimated on the basis of
single-species experiments and the literature.
Search Image for effects in more complex systems
The second aim of this project is to develop a search image for mild
forms of toxic stress in communities, and use this to recognize
effects of toxicants under field considitions. Functional aspects
(i.e. nutrient recycling) depend on structural aspects (biomass
amounts in the various trophic levels), but how mild toxic effects
will show up in both aspects is presently not known. Complex feedback
mechanisms in these integrated systems make application of a modeling
framework essential. We also want to determine the most relevant
single-species effects, to supplement and interpret measurements in
the field.
Food chains
The third aim of this project is to extend the search image for
effects on communities by accumulation of toxicants in food chains,
coupled to effects, and to apply our search image for effects in
communities to recognize effects of toxicants in field data. The
species here involve fish and bentic fauna. Most parameters will be
obtained from the literatire, but we will set up a limited
experimental program to estimate key parameters from single-species
toxicity tests. This effort will be started in 2006.
| 11:45 | welcome by
Prof. Dr. Nico van Straalen
(Rector Magnificus) |
| promotor Prof. Dr. Bas Kooijman;
copromotor Dr. Bob Kooi |
| 11:45 | Introduction by Daniel |
| 11:55 | Dr. Markus Liebig |
ECT Oekotoxikologie, Flörsheim Main, Germany |
| 12:05 | Dr. Bert van Hattum |
Instituut voor Milieuvraagstukken, FALW, VU |
| 12:15 | Dr. Dick de Zwart |
RIVM, Bilthoven |
| 12:25 | Dr. Kees van Gestel |
Instituut voor Ecologische Wetenschappen, FALW, VU |
| 12:35 | Dr. Tjalling Jager |
Sectie Theoretische Biologie, FALW, VU |
| 12:45 | end of defence; start of closed meeting |
| 13:10 | ceremony |
| 13:20 | end of ceremony; lunch in the Basket |
Go to projects' entry