As shown in many natural ecosystems as well as in theoretical exercises, food webs can become unstable when the environment changes. For example, eutrophication with phosphorous has shifted many crystal clear lakes, dominated by water fleas and vascular plants, into muddy pools saturated with blue-green algae. But what exactly drives these changes in trophic structure? In order to make intelligible decisions in conservation biology, we need to fundamentally understand how ecosystems respond to environmental change.

 Strong trophic interactions are quite apparent. Aphids feeding on
phloem juices have a strong link with the plants they feed on. The
classic food chain (flower-wabbit-cat picture) comprises strong
interactions exclusively. As shown by the arrows, the herbivore can
only eat plants and the carnivore can only eat herbivores. However, if
the carnivore is replaced with an omnivorous species (such as in the
flower-wabbit-bear picture) there exist two links towards the highest
trophic level, one from the shared resource, and one from the
herbivore to the carnivore. If, for instance, only a small fraction of
the carnivore's food is from vegetable origin, then the link
between the plants and the omnivore species is said to be weak. Weak
links are omnipresent in nature, and therefore, if we are to
understand dynamics of natural food webs, we should also establish
their dynamic effects in model studies.
Strong trophic interactions are quite apparent. Aphids feeding on
phloem juices have a strong link with the plants they feed on. The
classic food chain (flower-wabbit-cat picture) comprises strong
interactions exclusively. As shown by the arrows, the herbivore can
only eat plants and the carnivore can only eat herbivores. However, if
the carnivore is replaced with an omnivorous species (such as in the
flower-wabbit-bear picture) there exist two links towards the highest
trophic level, one from the shared resource, and one from the
herbivore to the carnivore. If, for instance, only a small fraction of
the carnivore's food is from vegetable origin, then the link
between the plants and the omnivore species is said to be weak. Weak
links are omnipresent in nature, and therefore, if we are to
understand dynamics of natural food webs, we should also establish
their dynamic effects in model studies.
We studied the effect of an omnivorous link on the dynamics of a mathematical food web model, and compared the results obtained, to similar results derived from a model of a classic food chain. In this case, we used a microbial food web, living in a chemostat, where non-living resources are supplied at a constant rate, and all of the chemostat's constituents are continuously flushed out at a fixed rate to maintain a constant volume. The food web structure is as follows: a producer utilizes the non-living resources, a predator feeds upon producers and an omnivorous species eats predators, as well as producers. It appeared that indeed, the replacement of a top-predator by an omnivore reduced the food-web's potential to oscillate or act chaotically. Enrichment with resources (compare with eutrophication) caused the food web to become unstable, but especially when the omnivore was only weakly linked to the producer species, this effect was far less severe than in the food chain.
The presence of the many non-limiting nutrients in real ecosystems may impose weak interactions on food webs. Maybe, after all, we should not expect food webs to destabilize when we add more resources. Or maybe we need better models that deal with multiple nutrients in the environment, and establish the effects of increasing the particular resources.
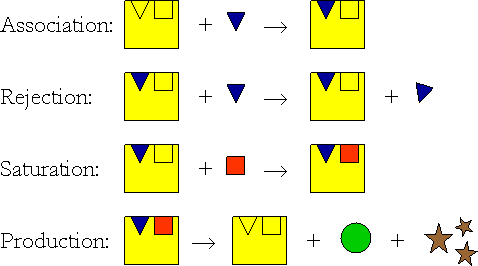 Traditional methods for modeling multiple nutrient limitation have
problems in their mathematical tractability, due to inconvenient
switches imposed on those models. In the late nineties, Bas Kooijman
proposed a way to circumvent these problems. He assumed that the
transformation from the required nutrients into metabolites or biomass
is analogous to enzymatic reactions, where substrates are transformed
into products. Underlying is the assumption that organisms require all
nutrients in fixed proportions for any given process. With slight
simplifications from the mathematical formulation of enzyme kinetics,
it appears to be possible to have simple equations for modeling
stoichiometry in a population/ecosystem context. The Synthesizing Unit
(SU) was born. The scheme of the SU shows how it works by taking a
very simple example. Suppose that the blue triangle and the red square
are packages of substrates, in just the quantities required for the
production of some compound. These substrates are, for instance,
reserves which are mobilized in the organism. At any time, the SU
notices a flux of squares and triangles, and it will pick them up in a
process called association. However, when the SU is saturated with
blue triangles, it can not bind another of those; it has to wait until
it binds a red square and if that happens, production starts. In this
case, a nice green circle is produced, but, as things are in nature,
this transformation will hardly be 100% efficient and some
by-products, indicated as brown stars appear as well. If there is an
excess of blue triangles coinciding with a lack of red squares,
production is slow. The production is fastest when all substrates are
present in the proper proportions.
Traditional methods for modeling multiple nutrient limitation have
problems in their mathematical tractability, due to inconvenient
switches imposed on those models. In the late nineties, Bas Kooijman
proposed a way to circumvent these problems. He assumed that the
transformation from the required nutrients into metabolites or biomass
is analogous to enzymatic reactions, where substrates are transformed
into products. Underlying is the assumption that organisms require all
nutrients in fixed proportions for any given process. With slight
simplifications from the mathematical formulation of enzyme kinetics,
it appears to be possible to have simple equations for modeling
stoichiometry in a population/ecosystem context. The Synthesizing Unit
(SU) was born. The scheme of the SU shows how it works by taking a
very simple example. Suppose that the blue triangle and the red square
are packages of substrates, in just the quantities required for the
production of some compound. These substrates are, for instance,
reserves which are mobilized in the organism. At any time, the SU
notices a flux of squares and triangles, and it will pick them up in a
process called association. However, when the SU is saturated with
blue triangles, it can not bind another of those; it has to wait until
it binds a red square and if that happens, production starts. In this
case, a nice green circle is produced, but, as things are in nature,
this transformation will hardly be 100% efficient and some
by-products, indicated as brown stars appear as well. If there is an
excess of blue triangles coinciding with a lack of red squares,
production is slow. The production is fastest when all substrates are
present in the proper proportions.
 The methods above make it possible to create a mathematical food web
model in which multiple nutrient limitation is dealt with in a
convenient way. We analyzed a model of a small food chain, where algae
use minerals of different kinds in order to grow, while grazers feed
on these algae. Meanwhile, thanks to realistic formulations of
biochemical transformations, unused by-products of any organism, as
well as detritus of deceased organisms, can be traced and recycled in
the environment, where micro-organisms reduce these compounds to
minerals that can be taken up by the algae again. We modeled a fully
functional marine ecosystem, including the copepod Acartia
tonsa and the diatom Thalassiosira weissfloggii, in this
way (see the food-web diagram), and parameterized it using literature
data. The model physiologically based model of Acartia tonsa was
created in cooperation with Tom
Anderson from the Southampton Oceanography Center. We ended up
with a number of surprising conclusions. First of all, the food web
destabilized at high levels of nitrogen-enrichment, however, if carbon
was added in excess to the food web, it remained stable. If carbon
however was depleted, the food-web destabilized again. This suggests
that a proper relative availability of potentially limiting compounds
is important to food web stability, and that the classic "paradox of
enrichment" is only an exponent of that. Furthermore, we could see
that nutrient limitation on the lowest trophic level did not
necessarily lead to limitation at higher trophic levels. Apparently,
looking at food webs in a more nutritionally detailed manner enables
us to fine-tune our classic ideas on nutrient enrichment and
limitation.
The methods above make it possible to create a mathematical food web
model in which multiple nutrient limitation is dealt with in a
convenient way. We analyzed a model of a small food chain, where algae
use minerals of different kinds in order to grow, while grazers feed
on these algae. Meanwhile, thanks to realistic formulations of
biochemical transformations, unused by-products of any organism, as
well as detritus of deceased organisms, can be traced and recycled in
the environment, where micro-organisms reduce these compounds to
minerals that can be taken up by the algae again. We modeled a fully
functional marine ecosystem, including the copepod Acartia
tonsa and the diatom Thalassiosira weissfloggii, in this
way (see the food-web diagram), and parameterized it using literature
data. The model physiologically based model of Acartia tonsa was
created in cooperation with Tom
Anderson from the Southampton Oceanography Center. We ended up
with a number of surprising conclusions. First of all, the food web
destabilized at high levels of nitrogen-enrichment, however, if carbon
was added in excess to the food web, it remained stable. If carbon
however was depleted, the food-web destabilized again. This suggests
that a proper relative availability of potentially limiting compounds
is important to food web stability, and that the classic "paradox of
enrichment" is only an exponent of that. Furthermore, we could see
that nutrient limitation on the lowest trophic level did not
necessarily lead to limitation at higher trophic levels. Apparently,
looking at food webs in a more nutritionally detailed manner enables
us to fine-tune our classic ideas on nutrient enrichment and
limitation.
For more info on this project, see the Kuijper, 2004.